Active system
Enzyme-Substrate-Inhibitor System
Please use the default enzyme concentration [E], substrate concentration [S], Michaelis-Menten constant Km, and IC50 to convert to Ki value OR enter your own value.
CLASSIC INHIBITOR EXAMPLE (botulinum neurotoxin type A) - Ref Burnett et al. 2007
System Selection
References
OTHER EXAMPLES
- Tight-binding inhibitor (monoamine oxidase A) - Ref Fowler CJ and Strolin BM 1983
- Complex and potentially cooperative inhibitor (CYP3A4) - Ref Marechal JD et al. 2006
Links: Ki Calculator For Fluorescence-Based Competitive Binding Assays, University of Michigan
More Information
Background
Some analyses of networks, pathways and metagenomics focus on identifying key proteins or polynucleic acids as targets for inhibitory compounds. Typically, high throughput screening assays are used initially to compare and down-select potential inhibitors of enzymatic activity or macromolecule-ligand binding. Many functional assays seek a total inhibitor concentration that reduces these activities by 50% (IC50). However, the IC50 value depends on concentrations of the enzyme (or target molecule), the inhibitor, and the substrate (or ligand) along with other experimental conditions. What is required is an accurate determination of the Ki value, an intrinsic, thermo-dynamic quantity that is independent of the substrate (ligand) but depends on the enzyme (target) and inhibitor. Thus, comparisons can be more readily made among different laboratories to characterize the inhibitors. While these more time-consuming assays are usually done with the most promising candidates, accurate, initial estimates of Ki values would be beneficial. A much discussed problem in the literature (e.g.. (Brandt et al. 1987 Biochem Med Metab Biol 37:344) is converting IC50 to Ki values because even the simplest types of inhibitory mechanisms (competitive, uncompetitive, non-competitive) will influence the calculation. To help address this problem, our web server tool calculates Ki values from IC50 values using equations from the literature for enzyme-substrate and target-ligand interactions by different inhibitory mechanisms. Additional calculations are performed for tightly bound inhibitors of enzyme-substrate reactions in which free, rather than total, concentrations of the molecular species are calculated for non-classic Michaelis-Menten kinetics. Similar calculations can be performed for target molecule-ligand systems. This calculator will enable end users to judge the quality of the underlying assumptions for these calculations. User-defined input values include total concentrations of the enzyme (or target molecule) and substrate (or ligand), the Km of the enzyme-substrate (or the Kd of the target-ligand) reaction and the IC50 value. The outputs include tabulations of the Ki values under different kinetic schemes, extensive tabulations of the results and the corresponding equations. Help buttons are available for Background, Assumptions, Literature, Links and Equations along with examples taken from the IC50_Ki_Converter host database that contains kinetic information on neurotoxin inhibitors.Assumptions
It is assumed that all of the substrate and inhibitor binding reactions are reversible and that they all have a one-to-one stoichiometry, i.e., no multiple binding of inhibitor molecules or any form of cooperativity, or other complex mechanisms of inhibition such as partial or mixed types. It is also assumed that in the enzymatic reactions that enzyme autocleavage did not occur and that when substrates for fluorescence resonance energy transfer were used, appropriate corrections for inner filter effects were performed. Comparison of Km or IC50 values for a set of inhibitor candidates is only assumed to be valid when they are evaluated under identical experimental conditions. In most experimental studies of enzyme kinetics, the total concentrations of substrate and inhibitor used are in excess to make their free and total concentrations essentially the same. Under the conditions of some ligand-receptor (e.g., protein) binding studies, the free concentrations become sufficiently important to require modifications of these equations.Literature
- Munson P.J., Rodbard D. (1988). An exact correction to the "Cheng-Prusoff" correction. J Recept Res 8:533-46.
- Cheng Y., Prusoff W.H. (1973). Relationship between the inhibition constant (KI) and the concentration of inhibitor which causes 50 per cent inhibition (I50) of an enzymatic reaction. Biochem Pharmacol 22:3099-108.
- Brandt R.B., Laux J.E., Yates S.W. (1987). Calculation of inhibitor Ki and inhibitor type from the concentration of inhibitor for 50% inhibition for Michaelis-Menten enzymes. Biochem Med Metab Biol 37:344-9.
- Lazareno S., Birdsall N.J. (1993). Estimation of competitive antagonist affinity from functional inhibition curves using the Gaddum, Schild and Cheng-Prusoff equations. Br J Pharmacol 109:1110-9.
- Henderson, P.J. (1972) A linear equation that describes the steady-state kinetics of enzymes and subcellular particles interacting with tightly bound inhibitors. Biochem. J., 127:321-333.
- Copeland, R.A., Lombardo, D., Giannaras, J., Decicco, C.P. (1995) Estimating KI values for tight binding inhibitors from dose-response plots Bioorg. Med. Chem. Lett., 5:1947-1952.
- Cheng H.C. (2001). The power issue: determination of KB or Ki from IC50. A closer look at the Cheng-Prusoff equation, the Schild plot and related power equations. J Pharmacol Toxicol Methods 46:61-71.
- Huang X. (2003). Fluorescence polarization competition assay: the range of resolvable inhibitor potency is limited by the affinity of the fluorescent ligand. J Biomol Screen 8:34-8.
- Nikolovska-Coleska Z., Wang R., Fang X., Pan H., Tomita Y., Li P., Roller P.P., Krajewski K., Saito N.G., Stuckey J.A., Wang S. (2004). Development and optimization of a binding assay for the XIAP BIR3 domain using fluorescence polarization. Anal Biochem 332:261-73.
- Lebeda, F.J., Adler, M., Erickson, K., Chushak, Y. (2008) Onset dynamics of type A botulinum neurotoxin-induced paralysis. J. Pharmacokinet. Pharmacodyn., 35:251-67.
- Craig, D.A. (1993) The Cheng-Prusoff relationship: something lost in the translation. Trends Pharmacol. Sci., 14:89-91.
- Mudunuri U., Stephens R., Bruining D., Liu D. and Lebeda F.J. (2006). botXminer: mining biomedical literature with a new web-based application. Nucleic Acids Research 34:W748-W752.
- Barthelmes, J., Ebeling, C., Chang, A., Schomburg, I., Schomburg, D. (2007) BRENDA, AMENDA and FRENDA: the enzyme information system in 2007. Nucleic Acids Res., 35:D511-D514.
- Greene, J.M., Collins, F., Lefkowitz, E.J., Roos, D., Scheuermann, R.H., Sobral, B., Stevens, R., White, O., Di Francesco, V. (2007) National Institute of Allergy and Infectious Diseases bioinformatics resource centers: new assets for pathogen informatics. Infect. Immun., 75:3212-3219.
- Berman, H.M., Westbrook, J.D., Gabanyi, M.J., Tao, W., Shah, R., Kouranov, A., Schwede, T., Arnold, K., Kiefer, F., Bordoli, L., Kopp, J., Podvinec, M., Adams, P.D., Carter, L.G., Minor, W., Nair, R., La Baer, J. (2009) The protein structure initiative structural genomics knowledgebase. Nucleic Acids Res., 37:D365-D368.
- Fowler, C.J., Strolin Benedetti, M. (1983) Cimoxatone is a reversible tight-binding inhibitor of the A form of rat brain monoamine oxidase. J. Neurochem., 40:510-513.
- Burnett, J.C., Opsenica, D., Sriraghavan, K., Panchal, R.G., Ruthel, G., Hermone, A.R., Nguyen, T.L., Kenny, T.A., Lane, D.J., McGrath, C.F., Schmidt, J.J., Vennerstrom, J.L., Gussio, R., Solaja, B.A., Bavari, S. (2007) A refined pharmacophore identifies potent 4-amino-7-chloroquinoline-based inhibitors of the botulinum neurotoxin serotype A metalloprotease. J. Med. Chem., 50:2127-2136.
- Marechal, J.D., Yu, J., Brown, S., Kapelioukh, I., Rankin, E.M., Wolf, C.R., Roberts, G.C., Paine, M.J., Sutcliffe, M.J. (2006) In silico and in vitro screening for inhibition of cytochrome P450 CYP3A4 by comedications commonly used by patients with cancer. Drug Metab. Dispos., 34:534-8.
- Cheng H.C. (2004). The influence of cooperativity on the determination of dissociation constants: examination of the Cheng-Prusoff equation, the Scatchard analysis, the Schild analysis and related power equations. Pharmacol Res. 2004 Jul;50(1):21-40.
Reactions
Competitive Inhibition
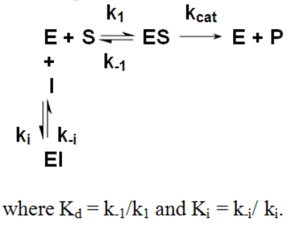
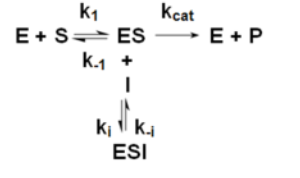
Uncompetitive Inhibition
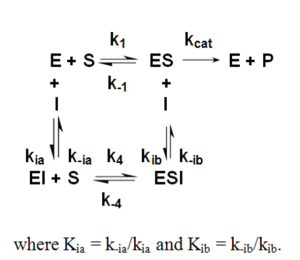
Noncompetitive Inhibition
Michaelis-Menten Enzyme-Substrate-Inhibitor Interaction
Competitive InhibitionKi= IC50/(S/Km+1)
Uncompetitive Inhibition
Ki= IC50/(Km/S+1)
Noncompetitive Inhibition
Ki= IC50
Michaelis-Menten Enzyme-Substrate-Inhibitor Interaction for Tightly Bound Inhibitors
Competitive InhibitionKi= (IC50- E/2)/(S/Km+1)
Uncompetitive Inhibition
Ki= (IC50- E/2)/(Km/S+1)
Noncompetitive Inhibition
Ki= IC50- E/2
Protein-Ligand-Inhibitor Interaction
Competitive Species
Ki = I50/[(L50/Kd) + (P0/Kd)+1][P0], free protein at 0% inhibition (positive control)
P0 = [-(Kd+L-P)+((Kd+L-P)2+4PKd)0.5]/2
[L0], free ligand at 0% inhibition (positive control)
L0 = L - PL0
[PL0], protein-ligand complex at 0% inhibition (positive control)
PL0 = P-P0
[P50], free protein at 50% inhibition
P50 = Kd * PL50/L50
[L50], free ligand at 50% inhibition
L50 = L-PL50
[PL50]. protein-ligand complex at 50% inhibition
PL50 = PL0/2
[I50], free inhibitor at 50% inhibition
I50 = IC50 - P +PL50[1+(Kd/L50)]
[PLI50], protein-inhibitor complex at 50% inhibition
PLI50 = IC50 - I50
Noncompetitive Species
Ki = I50/[(2P0/(P-P0))-(Kd/L50)+1][P0], free protein at 0% inhibition (positive control)
P0 = [-(Kd+L-P)+((Kd+L-P)2+4PKd)0.5]/2
[L0], free ligand at 0% inhibition (positive control)
L0 = L - PL0
[PL0], protein-ligand complex at 0% inhibition (positive control)
PL0 = P-P0
[P50], free protein at 50% inhibition
P50 = Kd * PL50/L50
[L50], free ligand at 50% inhibition
L50 = [-(P-L)+((P-L)2-4(-PL0Kd/2))0.5]/2
[PL50]. protein-ligand complex at 50% inhibition
PL50 = PL0/2
[I50], free inhibitor at 50% inhibition
I50 = IC50 - P+(PL0/2)(1+(Kd/L50))
[PLI50], protein-ligand-inhibitor complex at 50% inhibition
PLI50 = P - PL50 - P50